http://93.174.130.82/news/shownews.aspx?id=7d5310b0-8c9c-4f45-9e25-102688342779&print=1
© 2025 Российская академия наук
Довольно давно стало очевидно, что исследование биологического разнообразия какого-либо региона не ограничивается выявлением только числа обитающих там видов. Оказалось, что для многих групп живых организмов характерны и внутривидовые морфо-экологические формы. По этой причине реальный уровень локального разнообразия может быть намного выше, чем описываемый только числом видов. Одними из лидеров в этом являются рыбы, для ряда видов которых характерен высокий уровень внутривидовой морфологической и экологической изменчивости, когда образуются внутривидовые морфотипы и экологические формы.
Такая внутривидовая диверсификация характерна для многих лососёвых (Salmonidae), сиговых (Coregonidae) и карпообразных (Cypriniformes) рыб. Для карпообразных хорошо известны пучки (флоки) форм мелких азиатских усачей рода Puntius из оз. Ланао на о-ве Минданао (Филиппины), а также центральноазиатских алтайских османов рода Oreoleuciscus (рис. 1) и крупных африканских усачей рода Barbus (=Labeobarbus) (рис. 2). Последние две группы рыб учёные Института проблем экологии и эволюции им. А.Н. Северцова РАН и Института биологии развития им. Н.К. Кольцова РАН уже много лет изучают в рамках работ Совместной Российско-Монгольской комплексной биологической экспедиции РАН и Монгольской академии наук и Российско-эфиопской биологической экспедиции РАН.

Материал собирали в двух «центрах формообразования рыб»: в Центрально-азиатском бессточном бассейне в западной Монголии и в водоемах Абиссинского нагорья (Эфиопия). Целью исследования был анализ сходства механизмов формирования различий в строении черепа африканских усачей комплекса Barbus intermedius и алтайских османов рода Oreoleuciscus в полиморфных и в мономорфных популяциях водоёмов Эфиопии и Монголии. Одна из задач заключалась в поиске индикатора интенсивности формообразования в разных популяциях. Было установлено, что в анализе главных компонент таким индикатором может служить наличие так называемого «челюстного кластера» в векторных нагрузках параметров каждой из трёх костей челюстной дуги (praemaxillare, maxillare, dentale) и функционально связанной с ними кости жаберной крышки interoperculum. Эти кости отвечают за величину максимального открытия рта.

Установлено, что в популяции усачей африканского озера Тана, где наряду со всеядными генерализованными особями, обитает ряд большеротых рыбоядных форм усачей, в выборке генерализованных особей «челюстной кластер» находится в векторных нагрузках на первую главную компоненту (ГК1), тогда как в мономорфных популяциях двух других африканских озёр: Аваса и Лангано «челюстной кластер» имеет место в векторных нагрузках на вторую главную компоненту (ГК2) (рис. 3).
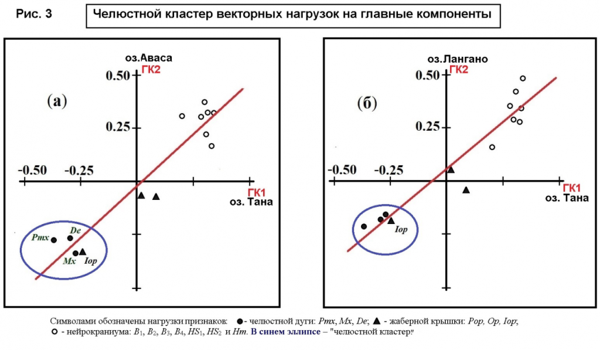
Это можно рассматривать как свидетельство того, что расположение «челюстного кластера» в ГК1 отражает высокую интенсивность формообразования, ведущую к появлению в водоёме внешне хорошо различимых специализированных форм, которые в случае оз. Тана одни авторы считают морфотипами, другие - придают им видовой статус. Расположение «челюстного кластера» в ГК2, судя по всему, свидетельствует о низкой интенсивности формообразования, недостаточной для возникновения внешне различимых форм. Вместе с тем, расположение «челюстного кластера» в ГК2, видимо, можно рассматривать, как некий потенциал возникновения специализированных форм от генерализованной при изменении условий обитания.
Для проверки такого предположения рассмотрим ситуацию в оз. Орог и впадающей в него р. Туин Центрально-Азиатского бессточного бассейна Монголии. Для оз. Орог характерно циклическое (в 12-13 лет) высыхание (Дгебуадзе, 2001). В «засушливый период», когда озеро высыхает, алтайские османы сохраняются только в р. Туин, где они представлены генерализованной речной формой, в выборке которой кластер векторных нагрузок признаков костей, формирующих челюсти, локализован в ГК2, подобно отмеченному выше кластеру в ГК2 генерализованных усачей эфиопских озёр Аваса и Лангано. Когда оз. Орог во «влажный период» наполняется водой, от скатившихся в него речных особей довольно скоро появляется именуемая «карликовой» озёрная генерализованная форма, у которой кластер признаков костей, формирующих челюсти, имеет место в векторных нагрузках уже не второй, а первой главной компоненты, как у генерализованных усачей оз. Тана. Далее от «карликовой» формы, для которой характерно питание растениями и беспозвоночными, возникает рыбоядная «озёрная" форма, по внешнему виду сходная с одной из рыбоядных форм африканских усачей оз. Тана (рис. 4).

Таким образом «челюстной кластер» действительно можно рассматривать как индикатор интенсивности формообразования в популяциях по крайне мере двух родов карповых рыб. И его обнаружение в векторных нагрузках на первую главную компоненту даёт весомые основания вести поиск специализированных форм в изучаемой популяции, даже если в первых пробах таковые обнаружены не были.
Полностью ознакомиться с результатами исследования можно в статье, опубликованной в Journal of Ichthyology.
Источник: ИПЭЭ РАН.