Cто лет назад было принято считать, что насекомые произошли от многоножек, а членистоногие в целом — от многощетинковых червей. За последние полвека было выдвинуто несколько гипотез, переворачивавших привычные взгляды. Популярным стало мнение о полифилии артропод. Молекулярные кладограммы подтвердили полузабытую идею о близости насекомых к ракообразным, но при этом объединили артропод не с кольчецами (в составе Articulata), а с круглыми червями (в составе Ecdysozoa). На основе синтеза данных морфологии, палеонтологии и генетики развития энтомологом Палеонтологического института им. А.А. Борисяка РАН предложен эволюционный сценарий, согласно которому высшие раки дали начало насекомым, многоножки возникли от шестиногих предков, первые панартроподы (динокариды) произошли от полихет, а не-артроподные группы экдизозоев (нематоды и прочие) испытали вторичное упрощение с утратой сегментации.
Для тех, кто в качестве отображения филогенетического родства между организмами приемлет только кладограмму, утверждение «насекомые родственны ракообразным» означает, что это две сестринские группы, произошедшие от неизвестных общих предков (не являющихся ни раками, ни насекомыми). На самом деле все гораздо интереснее. На самых примитивных насекомых — прыгающих щетинохвосток Archaeognatha (махилид и других) — больше всего похожи Syncarida (такие как тасманийские горные креветки), то есть достаточно продвинутые высшие раки. Это многостороннее сходство простирается вплоть до клеточных структур и характера экспрессии генов. Объяснить столь глубокое сходство конвергенцией невозможно, тем более что среда обитания у археогнат наземная, а у синкарид водная. Значит, насекомые — потомки высших раков (как предполагал еще в XIX в. немецкий зоолог Фриц Мюллер), и их предкам, прежде чем перейти на более высокий эволюционный уровень, пришлось преодолеть предыдущий, рачий.
Щетинохвосткам для прыжка за счет удара брюшком по субстрату на суше необходим больший объём брюшных мышц, чем креветкам для гребка хвостовым веером в воде. Поэтому при превращении раков в насекомых пять задних сегментов рачьей груди перестроились по образцу брюшных, так что брюшко стало длиннее, а число ног сократилось до трех пар. Подобные перестройки называют гомеотическими, как и гены, контролирующие рост и дифференцировку. Кроме того, насекомые унаследовали некоторые эмбриональные или личиночные черты синкаридных предков (такие изменения называют неотенией).
Забытая идея Эрнста Геккеля о том, что многоножки возникли от шестиногих предков, получила подтверждение со стороны генетики развития. У раков и насекомых подразделение туловища на грудь и брюшко регулируется одной и той же системой генов. Независимое приобретение двумя этими группами одинакового генетического механизма почти невероятно, зато его отключение могло происходить не один раз, приводя к превращению всех сегментов в грудные (снабженные ногами) и возникновению Myriapoda и подобных им форм (в том числе рачков Remipedia). Брюшко раков и примитивных насекомых обеспечивает экстренную локомоцию (гребок или прыжок). При скрытом образе жизни такая реакция бегства теряет смысл, поэтому многоножки и ремипедии утратили подразделение на грудь и брюшко и другие черты рачьих предков. Ногохвостки и их близкие родичи моделируют начальные этапы данного процесса («мириаподизации») у примитивных насекомых.
Таким образом, противоречие в вопросе о родственных связях насекомых — ближе они многоножкам или же ракообразным – мнимое, порожденное стремлением втиснуть родословное древо в прокрустово ложе дихотомий. Насекомые близки и высшим ракам, и многоножкам, но природа (полярность) этого родства различна. Насекомые возникли от высших раков (Malacostraca) и пошли по пути усложнения плана строения. Многоножки произошли от примитивных насекомых за счет упрощения плана строения.
«Многовесельные» рачки ремипедии и многоножки — отнюдь не близкие к исходным группам, как часто считают, а наоборот, высоко специализированные. Здесь уместна аналогия с безногими тетраподами. Подобно многоножкам, змееобразные наземные тетраподы в связи с ползанием в подстилке или роющим образом жизни вторично приобрели внешне гомономное туловище, содержащее более многочисленные метамеры (позвонки), и имеют головной отдел более специализированный, чем у их гетерономных предков. Подобно змеям, многоножки первично наземны, и нет причин предполагать для древнейших из них обитание в воде.
Подобно многоножкам, у змей весь осевой скелет видоизменен на манер груди — утрачены парные конечности, отсутствующие в исходном плане строения метамера позвоночных (см. ланцетника), и достигнуто это путем гомеотических изменений (области экспрессии генов, задающих развитие по типу груди, распространились на шейный и поясничный отделы). Так что по сути своей многоножки — это змеи среди членистоногих. И безногость змей, и «всеногость» многоножек — частичный возврат к исходному состоянию. Выводить щетинохвосток от многоножек не более логично, чем змей считать предками ящериц.
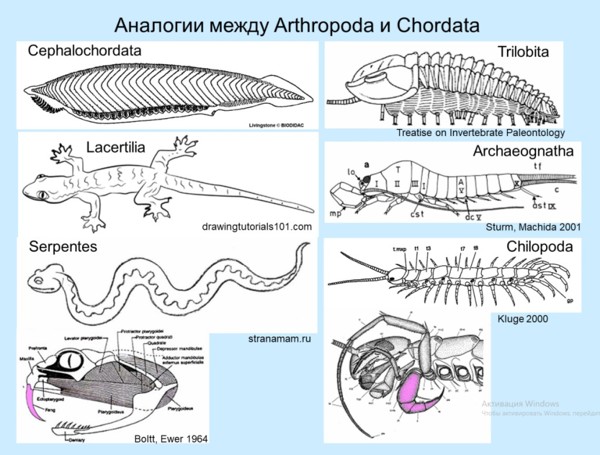
Аналогии между членистоногими и хордовыми
«Столбовую дорогу» эволюции членистоногих можно вслед за А.Г. Шаровым проследить вплоть до самых примитивных артропод (Dinocarida) раннего кембрия. Широко известный Anomalocaris и его родичи (Radiodonta) были первыми настоящими членистоногими, но членистыми у них были только хватательные или фильтрующие конечности на голове. Ходильные ноги впервые появились у «большеруких» (Megacheira), когда морское дно обогатилось кислородом и стало более пригодным для жизни.
К динокаридам относят и жаброносных лобопод (Opabinia и других), имевших под плавниками ходильные конечности, которые, как и головные, были кольчатыми (т.е. лобоподиями). От этих форм прямая дорога к ксенузиям, онихофорам и тихоходкам. Профессор Копенгагенского университета Клаус Нильсен предложил считать экдизозоев сестринской группой аннелид в составе артикулят, а круглых червей — утрачивающей сегментацию сестринской группой панартропод. Вероятно, противоречие «Articulata или Ecdysozoa» тоже лишь кажущееся: от ранних панартропод (жаброносных лобопод) возникли не только членистоногие, но и нематоды и родственные им группы, эволюционировавшие в сторону радикального упрощения плана строения. Ну а сами панартроподы произошли непосредственно от аннелид, точнее — от полихет.
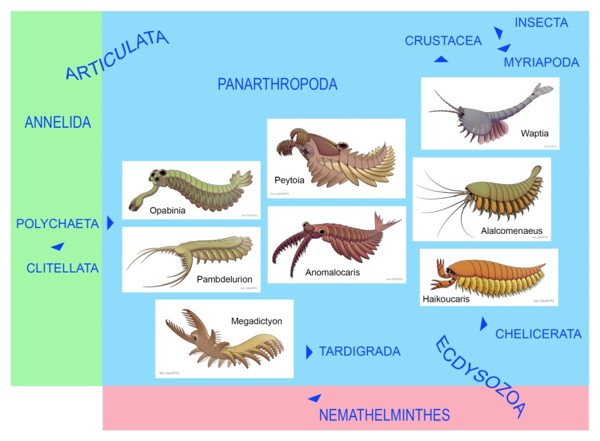
Некоторые кембрийские панартроподы в контексте филогении артикулят и экдизозоев: опабиния и памбделурион (жаброносные лобоподы), мегадиктион (ксенузии), пейтойя и аномалокарис (радиодонты), алалкоменеус и хайкоукарис (мегахейры), ваптия (крустацеоморфы)
Морфологическое сходство членистоногих с полихетами гораздо глубже, чем принято думать. Ближе всего к артроподам такие непохожие на червей существа, как морская мышь Aphrodita и другие чешуйчатые черви. Членистоногие могут быть неотеническими потомками таких полихет, утратившими хеты и выворачивающуюся глотку и сохранившими личиночное вентральное положение рта.
Хитин, у полихет присутствующий только в щетинках, челюстях и выстилке передней кишки, у панартропод стал основным компонентом покровов. Подобная экспансия морфогенетических механизмов от головы на все тело предполагается и в случаях возникновения плакоидной чешуи рыб, волосяного покрова млекопитающих и ходильных ног артропод.
Итак, кладистическая идеология неприложима к макроэволюции. Новые (дочерние) группы высокого ранга возникают от продвинутых, специализированных представителей предковой (материнской) группы в связи с проникновением в новую адаптивную зону и повышением уровня организации. Дочерние группы могут далее эволюционировать как путем прогрессивного совершенствования нового плана строения, так и путем его регрессивного упрощения.
Исследование опубликовано в Paleontological Journal.
Текст: Д.Е. Щербаков.
Источник: ПИН РАН.